Какое сердце у аллигатора сколько камер
Крокодилы являются самыми высокоорганизованными среди современных рептилий. Для рептилии представители этого отряда весьма смышлённы. Например, известны случаи, когда крокодилы ударами хвоста сбивали с прибрежных деревьев птенцов. Более того, крокодил поддаётся примитивной дрессировке и способен узнавать своего хозяина и подплывать на зов. Самки крокодилов – самые заботливые матери среди рептилий. Рептилиям не свойственна забота о потомстве, после его вылупления из яиц, хотя и встречаются исключения, например у некоторых видов сцинков. Однако их забота о своих детёнышах не идёт ни в какое сравнение с заботой крокодилицы о своём потомстве. Крокодилица бережно охраняет свою кладку и после вылупления малышей, долгое время всячески опекает их. Более того, иногда она помогает крокодилятам вылупиться из яйца, аккуратно перебирая яйцо в своей могучей пасти. Обычно же, детёныши справляются с этой задачей сами – для этой цели у них на кончике носа есть специальный роговой вырост, которым они пробивают скорлупу яйца при вылуплении. Своих детёнышей самка крокодила переносит в пасти. Порой бывает так, что другие самки усыновляют чужих детёнышей.
Маленькие крокодильчики окрашены ярче, чем взрослые особи. Первое время они очень уязвимы и их всюду поджидает опасность: на них нападают крупные рыбы, вараны, цапли, аисты, орланы и другие птицы. Другие, более крупные крокодилы, для них тоже представляют довольно серьёзную опасность, так как среди крокодилов практикуется каннибализм.
Крокодилы обладают очень прочным черепом с мощными челюстями (за исключением некоторых специализированных видов, таких как к примеру гавиал), благодаря которым их хватка крайне сильна. Однако мускулы открывающие пасть довольно слабые. У крокодилов подвижна не только нижняя, но и верхняя челюсть. Челюсти способны совершать только строго вертикальные движения. У представителей семейства аллигаторов (Alligatoridae) челюсти относительно шире и сильнее, чем у настоящих крокодилов (Crocodylidae). У аллигаторов при смыкании пасти зубной ряд верхней челюсти заходит за зубной ряд нижней, образуя таким образом ножницеобразный прикус, и при закрытой пасти у аллигаторов видны только зубы верхней челюсти. У настоящих же крокодилов зубы верхней челюсти входят в промежутки между зубами нижней челюсти, и при закрытой пасти у них виды как верхниии, так и нижние зубы. Некоторые специализированные в сторону рыбоядства формы семейства настоящих крокодилов, в частности африканский узкорылый крокодил (Crocodylus cataphractus), приобрели черты гавиалов (семейство Gavialidae) – длинную узкую морду с относительно тонкими и длинными зубами. Болотный крокодил (Crocodylus palustris) и тупорылый крокодил (Osteolaemus tetraspis), напротив, своей относительно широкой мордой напоминают аллигаторов.
Зубы крокодилов прочно сидят в специальных ячейках (альвеолах). Все они имеют одинаковую коническую форму и различаются только по величине. Основания зубов у крокодилов полые, в которых развиваются новые зубы, которые при случае замещают выпавшие. Таким образом, смена зубов у крокодилов постоянна на протяжении всей жизни. Зубы слабо загнуты назад, что препятствует крокодилам вырывать куски мяса из тела жертвы. Однако обладая сильной хваткой, крокодилы вцепившись в жертву, начинают вращаться вокруг своей оси и таким образом вырывают куски мяса у своей добычи. Иногда пара или более крокодилов помогают друг другу: в то время как один из них держит добычу, другой, вращаясь, вырывает из неё куски.
Хоаны у крокодилов перемещены назад, в глубину зева. Такая особенность позволяет крокодилам поедать свою жертву в воде без риска захлебнуться, а также дышать через ноздри, когда ротовая полость заполнена водой. У крокодила нет слюнных желез, но так как он поедает свою добычу в воде, это не является для него проблемой.
Кожа у крокодилов прочная. Тело сверху покрыто крепким панцирем, который образован остеодермами, покрытыми роговыми щитками. На голове остеодермы срастаются с костями черепа.
Основным двигателем в воде крокодилам служит их длинный и очень мускулистый хвост. Лапы во время плавания выполняют лишь вспомогательную функцию. Пальцы у крокодилов соеденены перепонкой, причём на передних лапах перепонки развиты слабо и едва заметны. Перепонки имеются только на задних лапах. Задние конечности длиннее и сильнее, чем передние, что помогает крокодилу делать резкий бросок во время нападения на жертву. Во время быстрого плавания крокодилы плотно прижимают все четыре конечности к туловищу и плывут, изгибаясь в горизонтальной плоскости засчёт своего мощного хвоста. Помимо двигательной функции в воде, хвост крокодилов выполняет роль дополнительного (помимо челюстей) оружия. Сильным ударом хвоста он может сбить свою жертву с ног или переломать ей ноги. Крокодилы – быстрые и маневренные пловцы, способные подолгу обходиться без кислорода (в неподвижном состоянии они могут обходиться без кислорода часами). На суше, кажужийся неповоротливым, крокодил способен развить довольно большую скорость для существа с такой комплекцией. Крокодил Джонстона (Crocodylus johnstoni), а также молодые крокодилы других видов способны даже передвигаться галопом (что не характерно для рептилий) и таким образом развивать до 20-ти км в час на которких дистанциях. Крокодилы обладают большой силой, но как и все рептилии, они не очень выносливы. Они не любят зря расходовать свою энергию и могут часами лежать в совершенно неподвижном состоянии. Они способны впадать в некое анабиозное состояние. Во время засушливого сезона нильские крокодилы (Crocodykus niloticus) вырывают глубокие норы и проводят там в таком состоянии сезон засухи.
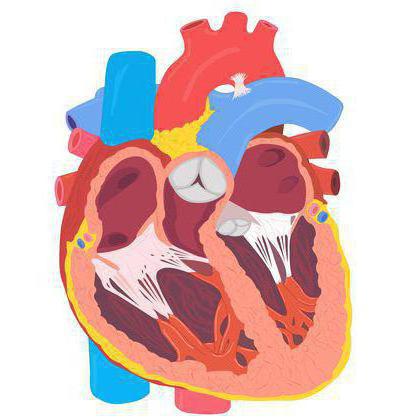
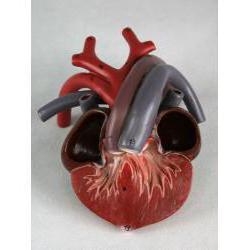
В отличие от других рептилий, крокодилы обладают четырёхкамерным сердцем (как и млекопитающие). Правый и левый желудочки разделены между собой перегородкой. Таким образом, кровь крокодилов разделена на венозную и артериальную. В левом желудочке она артериальная, в правом - венозная. Артериальная и венозная кровь перемешиваются при выходе, в месте слияния левой и правой дуг аорты.
Крокодилы – единственные из современных рептилий способные издавать рычащие звуки. Крокодилы обладают довольно неплохим слухом, однако плохо слышат высокие частоты. Во время брачного сезона самцы крокодилов испускают инфразвуковые сигналы, из-за которых вода на поверхности начинает "подпрыгивать". Зрение у крокодилов превосходное: они отлично видят в любое время суток как в водной среде, так и в воздушной. Зрачок у крокодилов имеет эллипсовидную форму. Третье веко очень хорошо развито и представляет собой прозрачную мембрану, которая защищает глаза крокодила во время плавания под водой.
Крокодилы являются одними из самых долгоживущих животных. Их продолжительность жизни примерно равна человеческой.
Иллюстрации:
Череп миссисипского аллигатора.
Череп молодого миссисипского аллигатора (вид сверху).
Череп молодого миссисипского аллигатора (вид снизу).
Челюсть миссисипского аллигатора.
В отличие от других рептилий, крокодилы обладают четырёхкамерным сердцем (как и млекопитающие).
А мне что-то помнится из школьного курса биологии, что сердце у них таки не вполне четырехкамерное: перегородка там есть, но на отдельные камеры она желудочек не разделяет. Не так?
Еще фото
Анатомия сердечно-сосудистой системы
У рептилий нет одной общей для всех схемы кровообращения. Однако можно выделить два основных типа строения сердца. Первый характерен для чешуйчатых и черепах, а второй – для крокодилов.
Ящерицы, змеи и черепахи
Сердце змей, ящериц и черепах трехкамерное, с двумя предсердиями и одним желудочком (Рис. 1-3). Такое строение предполагает возможность смешивания богатой кислородом крови из легких с кровью, ненасыщенной кислородом, идущей от систем органов. Ряд мышечных гребней и определенная периодичность сокращений служат для функционального разделения желудочка.
Правое предсердие получает ненасыщенную кислородом кровь, которая поступает от всех органов, через венозный синус – расширение на дорсальной стороне предсердия. Стенка венозного синуса мышечная, но не такая толстая, как стенка предсердия. Венозный синус получает кровь от четырех вен:
1. правой передней полой вены;
2. левой передней полой вены;
3. задней полой вены;
4. левой печеночной вены.
Левое предсердие получает насыщенную кислородом кровь из легких через левую и правую легочные вены.
В самом желудочке различают три полости: легочную, венозную и артериальную. Легочная полость – самый вентральный отдел, он продолжается краниально до устья легочной артерии. Артериальная и венозная полости расположены дорсальнее по отношению к легочной и получают кровь от левого и правого предсердия, соответственно. В своей самой краниальной и вентральной части венозная полость дает начало левой и правой дугам аорты (Рис. 4).
Мышечный гребень в некоторой степени отделяет легочную полость от других полостей. Артериальная и венозная полости объединены межжелудочковым каналом.
Одностворчатые атриовентрикулярные клапаны открываются с краниальной стороны межжелудочкового канала. Анатомически они организованы таким образом, что частично закрывают межжелудочковый канал во время систолы предсердия. Во время систолы желудочка их функция заключается в предотвращении регургитации крови из желудочка в предсердия. Серия мышечных сокращений и последующая разница давления в сердце рассматриваемых здесь рептилий разнесены по времени так, чтобы создать функционально двойную систему кровообращения. Систола предсердия нагнетает кровь в желудочек. Расположение атриовентрикулярных клапанов поперек межжелудочкового канала позволяет венозной крови из правого предсердия наполнять венозную и легочную полости. В то же время кровь из легких попадает из левого предсердия в артериальную полость. Желудочковая систола начинается с сокращения венозной полости. Последовательные сокращения венозной и легочной полостей выталкивают кровь из них в малый круг кровообращения с низким давлением.
В продолжение систолы сокращается артериальная полость, что приводит к движению крови через частично сокращенную венозную полость в большой круг кровообращения через левую и правую дуги аорты. Сокращение желудочка приводит мышечный гребень в положение непосредственной близости к вентральной стенке желудочка, создавая таким образом перегородку между артериальной и легочной полостями. Левый и правый атриовентрикулярные клапаны предотвращают возврат крови из желудочка в предсердия.
Все вышеупомянутые явления протекают только при нормальном дыхании. Такая система подачи крови ведет к ее сбросу слева направо на основе разницы давлений. При погружении под воду или в других ситуациях, когда легочное сопротивление и давление повышаются, движение крови происходит справа налево. У красноухой черепахи (Trachemys scripta elegans) при нормальном дыхании кровь движется преимущественно в легочном круге, который получает 60% объема выходящей из сердца крови, а оставшиеся 40% направляются ко всем системам органов. При погружении в воду кровь преимущественно движется по большому кругу, минуя легкие. В таких обстоятельствах давление в легочных сосудах выше, чем на периферии, поэтому кровь входит в сосуды с меньшим давлением – в дуги аорты. У ящериц кровь проходит в основном по левой дуге.
Крокодилы
Строение сердца у крокодилов очень напоминает таковое у птиц и млекопитающих, с той лишь разницей, что у крокодилов есть небольшое отверстие в межжелудочковой перегородке, разделяющей правый и левый желудочки – паниццево отверстие (foramen Pannizi), и что левая дуга аорты выходит из правого желудочка.
Строение сердца крокодилов двойственно по своей природе. Некоторое смешивание насыщенной и ненасыщенной кислородом крови может происходить через паниццево отверстие или в спинной аорте в месте слияния правой и левой дуг. Впрочем, при нормальном дыхании последнего варианта смешивания не происходит, так как давление в системном круге кровообращения превосходит давление в легочном круге. Сброс крови слева направо происходит через паниццево отверстие, и небольшое количество насыщенной кислородом крови попадает в правый желудочек.
Сам факт сброса крови справа налево при задержке дыхания и повышении сопротивления легочных сосудов может иметь большое клиническое значение. Рептилии под наркозом или без дыхания в отсутствии искусственной вентиляции легких могут демонстрировать непрогнозируемые реакции на ингаляционную анестезию. Кровообращение в обход легких может приводить к недостаточному распределению анестезирующих газов, таких как изофлюран, в системном круге для дальнейших манипуляций под анестезией. Значение длительного сброса справа налево, который может отмечаться при хронических воспалительных процессах в легких, до сих пор мало изучено. При этом можно ожидать серьезных изменений со стороны сердечно-сосудистой системы.
Воротная система почек
Воротная система почек – одна из частей венозной системы рептилий, вызывающая множество вопросов, потенциально имеющих значение для врача. Ее функция заключается в обеспечении достаточного кровоснабжения почечных канальцев при замедлении тока крови через клубочки для сохранения воды.
Афферентные вены воротной системы почек не проникают в клубочки; вместо этого они снабжают кровью проксимальные и дистальные извитые канальцы. Как и у млекопитающих, кровь к клеткам канальцев у рептилий подают приносящие артериолы, которые выходят из клубочков. Однако, в отличие от млекопитающих, в нефронах рептилий нет петель Генле и, следовательно, не происходит реабсорбции воды. В результате, для того чтобы сохранить воду, под воздействием аргининвазо-тоцина замедляется приносящий ток крови через клубочки. При снижении кровоснабжения в клубочках воротная система почек жизненно необходима для подачи крови к канальцам во избежание циркуляторного некроза.
Физиология сердечно-сосудистой системы
Частота сердцебиений у рептилий находится в довольно сложной зависимости от ряда факторов, в том числе температуры тела, размеров тела, уровня обмена веществ, дыхания и внешних раздражителей. Сердечная мышца характеризуется присущей ей максимальной производительностью, измеряемой максимальным напряжением сокращения в пределах зоны оптимальных предпочитаемых температур (ЗОПТ) для данного вида. В общем случае повышение активности приводит к повышению частоты сердцебиений. Частота может увеличиваться втрое по сравнению с частотой сокращений в состоянии покоя. Также, как правило, существует обратная зависимость между размером тела и частотой сердцебиений при заданной температуре.
Интересные вариации частоты сердцебиений при одной и той же температуре окружающей среды проявляются в зависимости от температурного статуса рептилии. В процессе нагревания животное обычно имеет более высокую частоту сердцебиений, чем во время охлаждения. Ускорение сердечного ритма при прогреве помогает достигнуть максимального поглощения тепла. Снижение частоты ударов сердца при понижении температуры окружающей среды помогает рептилии замедлить потерю тепла.
При низких температурах минутный объем сердца, по-видимому, поддерживается за счет увеличения его ударного объема. Ускоренное сердцебиение при повышенных температурах, очевидно, связано со скоростью обмена веществ. Теоретически, высокая частота сердцебиений должна ускорять транспорт кислорода. Изучение кислородного пульса (количество потребляемого кислорода при каждом сердцебиении в мл на массу тела в г) у разных видов говорит об отсутствии последовательной схемы взаимосвязи между объемом сокращения, поглощением кислорода и частотой сердцебиений ввиду повышенной потребности в кислороде при увеличении скорости метаболизма. Различные виды рептилий предположительно обладают множеством механизмов для улучшения подачи кислорода во время ускорения обмена веществ. Отдельно следует упомянуть тот факт, что скорость сердцебиений стремится к увеличению во время активного дыхания и уменьшению при задержке дыхания. Увеличение сердечного ритма совпадает со снижением легочного сопротивления и последующим увеличением легочного кровообращения. Соответственно, увеличение легочного кровообращения в период увеличения дыхательной активности служит для большей эффективности газообмена.
Сердечно-сосудистая система играет ключевую роль в терморегуляции рептилий. Как уже было сказано, частота ударов сердца повышается, когда животное нагревается, и понижается при его охлаждении. Несмотря на то что контролирующий механизм до конца не известен, изменения в кровеносной системе происходят до того, как изменится общая температура тела, что предполагает наличие кожных терморецепторов и барорецепторов.
При нагревании кожных покровов отмечается расширение сосудов в коже. Отток крови в периферические сосуды приводит к падению общего кровяного давления. Понижение сопротивления периферических сосудов способствует развитию сброса крови в сердце справа налево. Кровяное давление поддерживается, таким образом, на уровне, достаточном для снабжения кровью мозга и органов чувств по правой дуге аорты. Кроме того, так как кровь из кожных покровов возвращается в общее кровяное русло, повышается общая температура тела.
Снижение частоты сердцебиений при охлаждении кожи служит для сохранения тепла. При этом наблюдается сужение кровеносных сосудов в коже и относительное расширение сосудов в мышцах. Такое перераспределение крови призвано замедлить теплоотдачу.
Так же как для птиц и млекопитающих, изменения гемодинамики при погружении под воду очень важны и для рептилий. У них имеется ряд преимуществ по сравнению с теплокровными животными, так как рептилии могут использовать альтернативный путь метаболизма при отсутствии кислорода – анаэробный гликолиз. Способность выдерживать анаэробиоз различна у разных видов рептилий. Некоторые ящерицы выдерживают без кислорода не более 25 минут, тогда как некоторые виды черепах способны задерживать дыхание на 33 часа и более. Основные различия заключаются в разной толерантности миокарда к гипоксии.
Как правило, при погружении под воду развивается брадикардия. У крокодилов она обусловлена вагальным торможением сердца под некоторым влиянием торакального или внутрилегочного давления. При нырянии происходит симпатическое сужение кровеносных сосудов в скелетных мышцах, часто до ишемического порога. Такое повышение периферического сопротивления поддерживает кровяное давление для нормальной работы органов.
Сброс крови справа налево возникает при истощении запаса кислорода в паренхиме легких. При дальнейшем погружении сброс справа налево доминирует, практически полностью исключая подачу крови к легким. Общий сердечный выброс может снижаться до уровня в 5% по сравнению с нормальным состоянием. Способность снижать до минимума рабочую нагрузку на сердце, прокачивая лишь небольшую часть крови в системное русло, обеспечивает рептилиям явное преимущество при погружениях по сравнению с птицами и млекопитающими. Брадикардия, связанная с погружением, быстро обратима при первом же вдохе; у некоторых видов даже отмечено ускорение работы сердца еще до выхода на сушу.
Свойства кровеносной системы и их связь с газообменом на клеточном уровне должны приниматься во внимание в любых исследованиях в области кардиологии рептилий. Несмотря на кажущуюся несущественность данного вопроса, клинически подтверждено, что изменения в функционировании сердца или легких могут в значительной степени влиять на способность кровеносной системы к переносу кислорода и углекислого газа.
Молекула гемоглобина считается компонентом, от которого зависят респираторные свойства крови. Хотя структура гемоглобина рептилий пока полностью не описана, она, скорее всего, такая же, как у других позвоночных. Тем не менее, известен ряд существенных отличий в способности гемоглобина удерживать и отдавать кислород. Для этих отличий не было найдено каких-либо закономерностей в зависимости от условий среды, и они не являются общими для всего класса рептилий.
По мере растворения кислорода его давление (мера концентрации) приводит к насыщению или частичному насыщению гемоглобина. Молекула гемоглобина отвечает за респираторные свойства и цвет крови. Кривые диссоциации кислорода показывают, какое его количество удерживается гемоглобином при определенных условиях, и отражают влияние температуры, рН, углекислого газа, продуктов гликолиза, органических фосфатов в эритроцитах и таких ионов, как Na+, K+, Mg2+, Cl - , SO4 2- .
Если гемоглобин претерпевает изменения с момента рождения до формирования взрослой особи, то способность крови к насыщению кислородом будет различной в зависимости от этапа онтогенетического развития. При высокой скорости обмена веществ кривые диссоциации кислорода будут смещаться вправо, то есть сродство крови к кислороду будет ниже, что упрощает его доставку к тканям. У рептилий кривые диссоциации кислорода крайне вариабельны. Их сложно обобщить в связи с влиянием непостоянной температуры и скорости метаболизма, а также других перечисленных ранее факторов.
Разные рептилии обладают разными формами гемоглобина, и у некоторых видов гемоглобин эмбриона может иметь сходство к кислороду, отличное от такового у взрослых особей. Гемоглобин может по-разному принимать и отдавать кислород. Эти отличия часто не обнаруживаются клинически, но о них необходимо помнить, чтобы избежать излишней экстраполяции с одного вида на другой.
Сродство к кислороду является мерой того, насколько легко гемоглобин отдает кислород тканям. Гемоглобин с высоким сродством отдает кислород хуже. Низкое сродство означает лучшую отдачу кислорода. У рептилий обычно сродство гемоглобина к кислороду ниже, чем у млекопитающих. Эта адаптация позволяет снабжать кислородом ткани даже при небольшом его содержании в крови.
Во время нагрузок или стресса рептилии могут испытывать метаболический ацидоз вследствие образования молочной кислоты. Изменение рН крови снижает ее сродство к кислороду (эффект Бора), что приводит к тому, что кровь удерживает меньше кислорода и быстрее отдает его тканям.
Изучение кривых диссоциации кислорода у ряда видов рептилий не выявило для них определенных закономерностей. Однако можно предложить несколько общих концепций для отдельных групп рептилий.
Среди ящериц самые активные виды (например, те-йиды, веретеницевые) обладают, как и следовало ожидать, более низким сродством к кислороду. Более высокое сродство к кислороду характерно для медлительных рептилий или для хищников, поджидающих свою добычу (например, хамелеоны, гекконы). Некой серединой для сравнения можно считать игуановых (в том числе, Iguana iguana, Anolis spp., Ctenosaura spp.). Известно, что у игуановых ящериц сродство крови к кислороду напрямую связано с размерами тела. Однако данные, полученные путем измерений при предпочитаемой температуре, слишком ненадежны ввиду поведенческих различий между видами и потому не могут считаться клинически значимыми.
У черепах видимая разница существует между водными и сухопутными видами. Как правило, у водных видов сродство к кислороду ниже, то есть отдача кислорода происходит лучше. У некоторых черепах, живущих в условиях постоянной гипоксии, кровь обладает буферными свойствами, задерживающими эффект Бора, что можно считать адаптацией, связанной с необходимостью максимальной отдачи кислорода во время погружения. Неожиданным исключением является иловая красноватая черепаха (Kinosternum subrubrum), у которой кривая диссоциации кислорода такая же, как и у наземных черепах.
Змеи в этом вопросе принципиально отличаются от черепах. Сравнение водяной яванской бородавчатой змеи (Acrochordus javanicus) и обыкновенного удава (Constrictor constrictor) показало их противоположность по сродству к кислороду. У водяной змеи сродство к кислороду было выше, чем у наземной.
Эта разница может отчасти быть результатом усиленного эффекта Бора, отмечаемого у водных змей. Роль увеличения эффекта Бора, по-видимому, заключается в том, чтобы обеспечить доступность большего количества кислорода в периоды без дыхания при возрастании уровня CO2 в крови. Такая система насыщения крови кислородом позволяет этим видам отдавать кислород, когда это необходимо, во время погружения, и принимать кислород, когда он наиболее доступен, во время дыхательной вентиляции. У змей сродство к кислороду снижается с возрастом, тогда как кислородная емкость (процентный объем кислорода в полностью насыщенной крови) увеличивается по мере роста. Влияние размера тела на сродство к кислороду неодинаково; оно снижается с увеличением размеров (с возрастом) у змей, но повышается у ящериц.
Как и следовало ожидать, кислородная емкость достигает максимума, когда рептилия находится в зоне оптимальных предпочитаемых температур. У змей в связи с нерегулярным типом питания сродство к кислороду падает и его потребление резко возрастает во время переваривания пищи (процесса, требующего усиления обмена веществ). После приема большого количества пищи увеличивается не только потребление кислорода, но и размеры сердца. Андерсон и др. отмечают, что скорость обмена веществ после еды у тигрового питона (Python molurus bivitattus) может увеличиваться до 40%. Высокий уровень метаболизма может сохраняться до 14 дней.
Для поддержания такого уровня обмена сердце питона гипертрофируется на протяжении 48 часов после употребления пищи. Масса сердца может увеличиваться на 40% в ответ на увеличение экспрессии генов сократительных белков мышц. После завершения переваривания пищи размеры сердца возвращаются к норме.
В разных публикациях по этому вопросу приводятся различные данные. Состав сыворотки крови зависит не только от техники забора крови, но и от температуры внешней среды. Кроме того, он зависит от возраста животного. У 1 — 2-летних животных количество белка составляет 0,53 г/л, глюкозы — 4,9 ммоль/л; у 3 — 5-летних эти значения соответственно равны 0,61 г/ли 3,36 ммоль/л.
С помощью электрофореза можно наблюдать распределение белка. Ферограмма электрофореза сыворотки крови крокодилов показывает существенные отличия от распределения белка у млекопитающих в части соотношения альбумин/глобулин. Даже кайманы, крокодилы и аллигаторы отличаются в этом отношении друг от дpyra.
Частота сердцебиения, частота дыхания
В этой области проводилось множество исследовании. Показательны долговременные исследования. В изолированном помещении у кайманов в спокойном состоянии частот сердечных сокращений составляла 14 при 1,6 вдоха в минуту, в то время как у аллигаторов 12 сердечных сокращений при 0,6 вдоха. В присутствии человека значения утраивались. Кром того, на частоту сердечных сокращений и частоту дыхания оказывают влияние температура помещения и воды, а также время суток.
Кровяное давление
Систолическое давление у молодых иммобилизированных аллигаторов равняется 25 мм ртутного столба, диастолическое давление — 15 мм ртутного столба.
Температура тела
С помощью телеметрии были произведены длительные измерения температуры тела и оптимальной температуры у австралийского узкорылого крокодила и гребнистого крокодила, находившихся на свободе в большом парке. Значения для австралийского узкорылого крокодил колебались в пределах 25 — 36°С; оптимальной была температура 31,3 — 32,5°С. Для гребнистого крокодила значения температуры тела колебались в пределах 25,9 —35,5°С значение оптимальной температуры составляло 32,0 — 33,1°С. У аллигаторов были получены значения 32 —35°С, у только что убитых нильских крокодилов ректальна температура составляла 23 —29°С. У крокодилов температура тела не является вспомогательным средством для оценки состояния здоровья.
Содержание
Морфологические особенности
Длина большинства крокодилов 2—5,5 м. Их внешность демонстрирует адаптацию к обитанию в водной среде: голова плоская, с длинным рылом; туловище приплюснутое; хвост мощный, сжатый с боков; ноги довольно короткие. На передних конечностях — 5 пальцев, на задних — 4 (нет мизинца), соединённых перепонкой. Глаза с вертикально-щелевидным зрачком расположены очень высоко; ноздри и ушные отверстия под водой закрываются подвижными клапанами.
Разница между крокодилом и Кожные покровы
Кожа крокодилов покрыта прямоугольными роговыми щитками, которые на спине и животе располагаются правильными рядами. Под спинными и реже под брюшными щитками развиваются остеодермы (костные пластины), образующие панцирь. Остеодермы в дневное время аккумулируют тепло поступающее вместе с солнечным светом. Благодаря этому температура тела крокодила в течение суток колеблется в пределах одного градуса. Крокодила не следует отождествлять с теплокровными животными (птицами и млекопитающими), у которых постоянство температуры тела поддерживается за счёт собственного метаболизма.
Костная система
Череп диапсидный — с двумя динозавры и современные клювоголовые. Брюхо у крокодила защищено брюшными рёбрами, не связанными с позвоночником. Органы дыхания
Носоглоточный проход отделён от ротовой полости вторичным костным нёбом, что позволяет крокодилу держать под водой пасть открытой, продолжая дышать через выставленные над водой ноздри. При этом воду в дыхательное горло не пропускает особый клапан в глубине пасти (нёбная завеска), а воздух проходит в Кровеносная система
Кровеносная система крокодилов самая совершенная среди пресмыкающихся. Сердце четырёхкамерное: имеет два предсердия и два желудочка, полностью разделённых перегородкой. Однако у крокодилов сохранились обе дуги Органы пищеварения
Зубы у крокодила однотипные, конические, полые внутри — в этих полостях по мере снашивания развиваются новые зубы. Зубы в течение жизни крокодила меняются неоднократно. При этом зубы верхней и нижней челюстей совмещены таким образом, что против самых крупных зубов одной челюсти находятся самые мелкие зубы второй. Зубы сидят в Органы выделения и размножения
Мочевого пузыря нет, что, вероятно, связано с жизнью в воде. Излишки внутриполостной жидкости выводятся через Нервная система и органы чувств
По развитию головного мозга крокодилы стоят выше других пресмыкающихся: большие полушария головного мозга имеют кору. Из органов чувств у крокодилов наиболее развиты зрение и слух. На поверхности челюстей крокодила расположены многочисленные рецепторы давления, которые позволяют ему чувствовать вибрацию, исходящую от животных, находящихся в воде на большом расстоянии.
Образ жизни
Самка откладывает яйца в песок на отмели или зарывает в гнездо из грязи и гниющей листвы. Число яиц в кладке колеблется от 20 до 100. Самки ряда видов остаются возле кладки, охраняя яйца от врагов. Молодые крокодилы еще внутри яиц, ко времени вылупления издают квакающие звуки, и мать раскапывает кладку, помогая потомству выбраться. Все яйца вылупляются одновременно.
Крокодилы растут в течение всей жизни. Половозрелости они достигают в возрасте 8—10 лет; живут до 80—100 лет. Естественных врагов у взрослых крокодилов мало, но многие животные, особенно вараны, черепахи, птицы (марабу) и гиены, охотно поедают яйца и молодых крокодилов.
Эволюция
птицами, — единственные выжившие представители подкласса архозавров (Archosauria). Современные крокодилы являются остатками большой (до 20 семейств) группы крокодиловых, в позднем динозаврам и современным птицам, чем к другим рептилиям; ряд особенностей их организации позволяет считать их самыми высокоорганизованными из пресмыкающихся. Эволюция крокодилов шла в направлении наибольшего приспособления к водному образу жизни и хищничеству.
Значение для людей
Крокодилы в разной степени опасны для человека. Некоторые никогда не нападают на человека (гавиал), другие нападают систематически (гребнистый крокодил), третьи (нильский крокодил) нападают изредка. Мясо крокодилов съедобно и употребляется в пищу населением многих тропических стран. Кожа крокодилов, особенно аллигаторов, используется для изготовления различных галантерейных изделий (портфелей, чемоданов и т. п.). Хищническое истребление крокодилов привело к резкому сокращению их численности и принятию охранных мер. В ряде стран существуют специальные фермы по разведению крокодилов.
Крокодиловы слёзы
Классификация
Отряд крокодилов включает 3 семейства:
- Crocodylus acutusCrocodylus cataphractusCrocodylus intermedius
- Crocodylus johnstoni
- Crocodylus mindorensis
- Crocodylus moreletiiCrocodylus niloticus
- Crocodylus novaeguineae
- Crocodylus palustrisCrocodylus porosus
- Crocodylus rhombifer
- Crocodylus siamensis
-
Род Аллигаторы
- Род Кайманы
- Caiman crocodilus
- Caiman crocodilus crocodilus
- Caiman crocodilus fuscus
- Caiman crocodilus apaporiensis
См. также
Ссылки
Литература
-
// Богданов А. К. О Крокодилах в России. Очерки из истории заимствований и экзотизмов. — М.: НЛО, 2006, с. 146—181
Эта страница использует содержимое раздела Википедии на русском языке. Оригинальная статья находится по адресу: Крокодилы. Список первоначальных авторов статьи можно посмотреть в истории правок. Эта статья так же, как и статья, размещённая в Википедии, доступна на условиях CC-BY-SA .
![]()
Наша планета густо заселена животными различных классов, отрядов и видов. Ученые изучают их строение и функциональное значение отдельных органов. О том, какое сердце у земноводных и пресмыкающихся, читайте в статье.
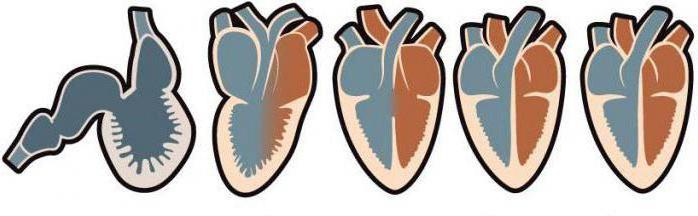
Как сердце из трех камер превратилось в четырехкамерное?
Позвоночные вышли на сушу из-за того, что их легочное дыхание стало интенсивно развиваться. Кровеносная система начала перестраиваться. Рыбы, дышащие жабрами, обладают одним кругом обращения крови, их сердце состоит всего из двух камер. Они не могут жить на суше.
![Четырехкамерное сердце имеют]()
Трех- или четырехкамерное сердце имеют наземные позвоночные. Они отличаются наличием двух кругов кровообращения. Их постоянная среда обитания – суша. Орган с тремя камерами имеют амфибии и рептилии. Хотя у отдельных видов пресмыкающихся имеется неполное его разделение на четыре части. Развитие настоящего четырехкамерного сердца в процессе эволюции происходило параллельно у млекопитающих, птиц и крокодилов.
Пресмыкающиеся и земноводные
У этих двух классов животных имеется по два круга обращения крови и сердце, состоящее из трех камер. Только у одной рептилии есть неполноценное, но обладающее четырьмя камерами, сердце. Это крокодил. Полноценный сердечный орган впервые появился у примитивных млекопитающих. В будущем сердце с таким строением унаследовали потомки динозавров – птицы. Оно передалось по наследству и современным млекопитающим.
Птицы
Четырехкамерное сердце имеют пернатые. Птицы отличаются полным разобщением кругов кровообращения: большого и малого, как у человека, когда не происходит смешивание крови – артериальной и венозной. Правая и левая половины органа полностью разделены.
У птиц четырехкамерное сердце, его строение представлено двумя предсердиями и таким же количеством желудочков. В желудочек венозная кровь поступает через правое предсердие. От него происходит отхождение легочной артерии, которая делится на левую и правую ветви. В результате кровь венозная оказывается в соответствующем легком. В это время кровь в легких окисляется и поступает в левое предсердие. Такое кровообращение называется его малым кругом.
![У птиц четырехкамерное сердце]()
Большой круг обращения крови берет начало с левого желудочка. От него отходит один-единственный сосуд, который называется правой дугой аорты, которая сразу на выходе из сердца отделяет две безымянные артерии: левую и правую. Сама же аорта разворачивается в области расположения правого бронха и идет параллельно позвоночному столбу уже в качестве спинной аорты. Каждая безымянная артерия разделяется на сонную и подключичную. Первая идет в голову, а вторая снова разделяется на грудную и плечевую. От спинной аорты отходят крупные артерии. Непарные предназначены для снабжения кровью желудка и кишечника, а парные – задних конечностей, органов полости таза и мышц стенок брюшины.
Четырехкамерное сердце имеют птицы, оно отличается тем, что у пернатых движение крови осуществляется в основном по сосудам крупных размеров, и только небольшая ее часть поступает в почечные капилляры. Птицы отличаются наличием крупного сердца с частыми сокращениями и поступлением в органы только чистой артериальной крови. Это позволило считать птиц теплокровными животными.
Кровеносная система млекопитающих
У млекопитающих четырехкамерное сердце, как у человека или птиц. Его формирование с полным разделением кругов обращения крови вызвано необходимостью развития такого качества, как теплокровность. Это объясняется так: теплокровные животные испытывают постоянную потребность в кислороде, удовлетворить которую способна лишь чистая кровь артерий с большим количеством кислорода. Обеспечить ею организм способно только четырехкамерное сердце. А смешанная кровь позвоночных, у которых сердце имеет три камеры, не способна дать нужную температуру тела. Поэтому такие животные и называются хладнокровными.
![У млекопитающих четырехкамерное сердце]()
Благодаря наличию полных перегородок кровь не смешивается. По большому кругу обращения течет только артериальная кровь, которой в нужной мере снабжаются все органы млекопитающего, что способствует ускорению обмена веществ. Этот процесс способствует поддержанию температуры на постоянном уровне. Четырехкамерное сердце имеют млекопитающие, птицы и другие классы животных, которым жизненно необходима постоянная и устойчивая температура тела. Теперь окружающая среда не влияет на них.
Ящерицы
На самом деле сердце у этих пресмыкающихся имеет три камеры с двумя предсердиями и одним желудочком. Но принцип его работы дает возможность утверждать, что четырехкамерное сердце имеют ящерицы. Объяснение это явление имеет следующее. Венозная полость заполняется бедной кислородом кровью, источником поступления которой является правое предсердие. Артериальная кровь, обогащенная кислородом, поступает из противоположного предсердия.
![Четырехкамерное сердце имеют ящерицы]()
Легочная артерия и обе дуги аорты сообщаются. Казалось бы, кровь должна полностью смешаться. Но этого не происходит, так как наличие мышечного лоскута в совокупности с двухфазным сокращением желудочка и дальнейшая работа сердца препятствуют смешиванию крови. Оно имеется, но в очень маленьких количествах. Поэтому по функциональному значению трехкамерное сердце ящериц похоже на четырехкамерное.
Рептилии
Крокодил имеет четырехкамерное сердце, хотя круги обращения крови полностью не разделены перегородкой. У пресмыкающегося орган (сердце), отвечающий за снабжение всего организма питанием через кровь, имеет особое строение. Кроме легочной артерии, отходящей от желудочка с правой стороны, имеется дополнительная, левая. По ней основная масса крови поступает в пищеварительную систему.
![Крокодил имеет четырехкамерное сердце]()
Между двумя артериями, правой и левой, сердце крокодила имеет отверстие. Через него кровь из вен имеет возможность попадать в большой круг обращения, и наоборот. Ученые долго считали, что сердце рептилии имеет тип переходного характера на пути следования к развитию полноценного сердца из четырех камер, как у теплокровных млекопитающих. Но это не так.
Черепахи
Система сосудов и сердца у этих пресмыкающихся такая же, как у других рептилий: сердце с тремя камерами, соединенные между собой вены и артерии. Содержание недостаточно окисленной крови увеличивается, когда возрастает внешнее давление. Это может происходить, когда животное ныряет или быстро передвигается. Частота сокращений сердца уменьшается, хотя значительно увеличивается концентрация углекислого газа.
![Четырехкамерное сердце имеют черепахи]()
Четырехкамерное сердце имеют черепахи, хотя по физиологическому строению орган имеет всего три камеры. Дело в том, что сердце черепахи отличается неполной перегородкой желудочка, вокруг которой кровь функционирует, имея разное количество кислорода.
Читайте также:
- Caiman crocodilus
-
Alligator mississippiensisAlligator sinensis